An important
step in understanding new scientific concepts is learning the
scales of the problem. How fast do processes occur? Over what
spatial scales? How much energy is consumed? In our courses we
always begin by looking at various cells and organisms to discern
the overall size, sizes of organelles, and rates of whole-cell
and intracellular movement, using a variety of light and fluorescence
microscopy techniques.
First,
students learn how to use bright-field microscopy to calibrate
different
magnifications on the microscope, enabling them to relate what
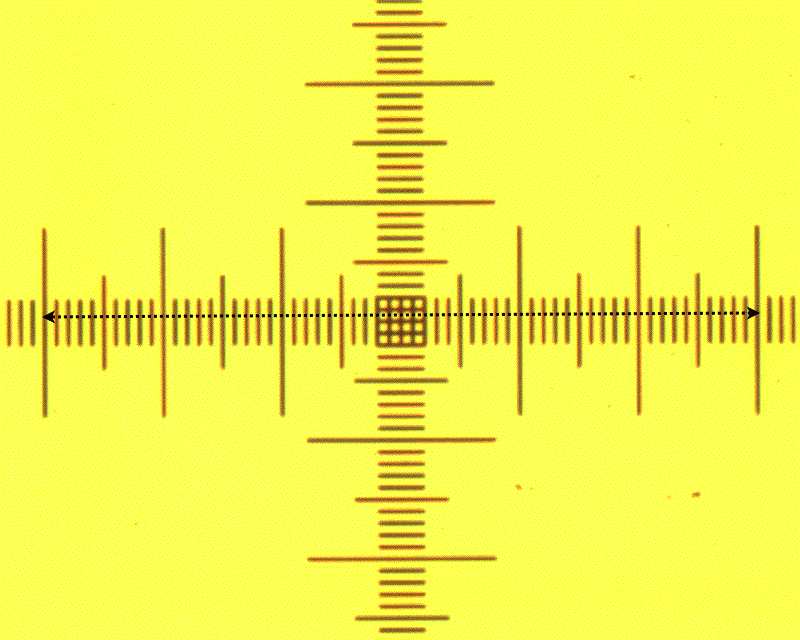
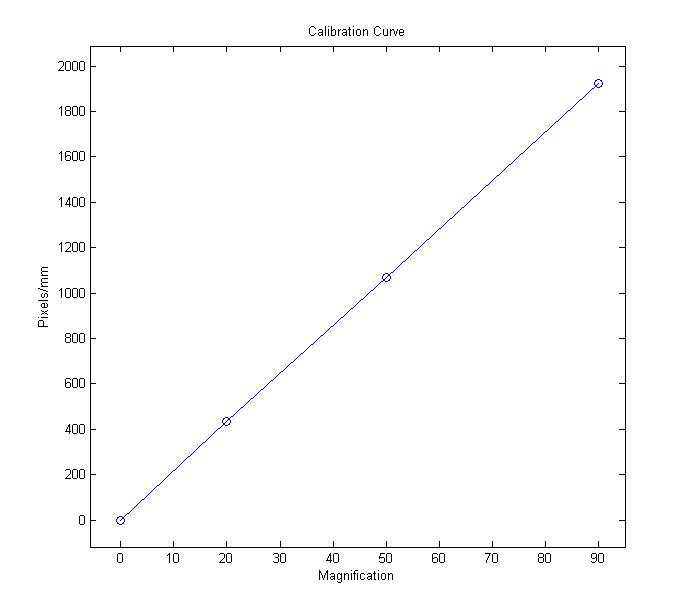
they see in the microscope to physical distances. A lithographic
graticule with 10um markers is imaged at different magnifications
and a calibration curve is generated as shown below.
While absolute
units of measurement are supremely useful, there is some merit
in simply having a 'feel' for the scale of the objects one is
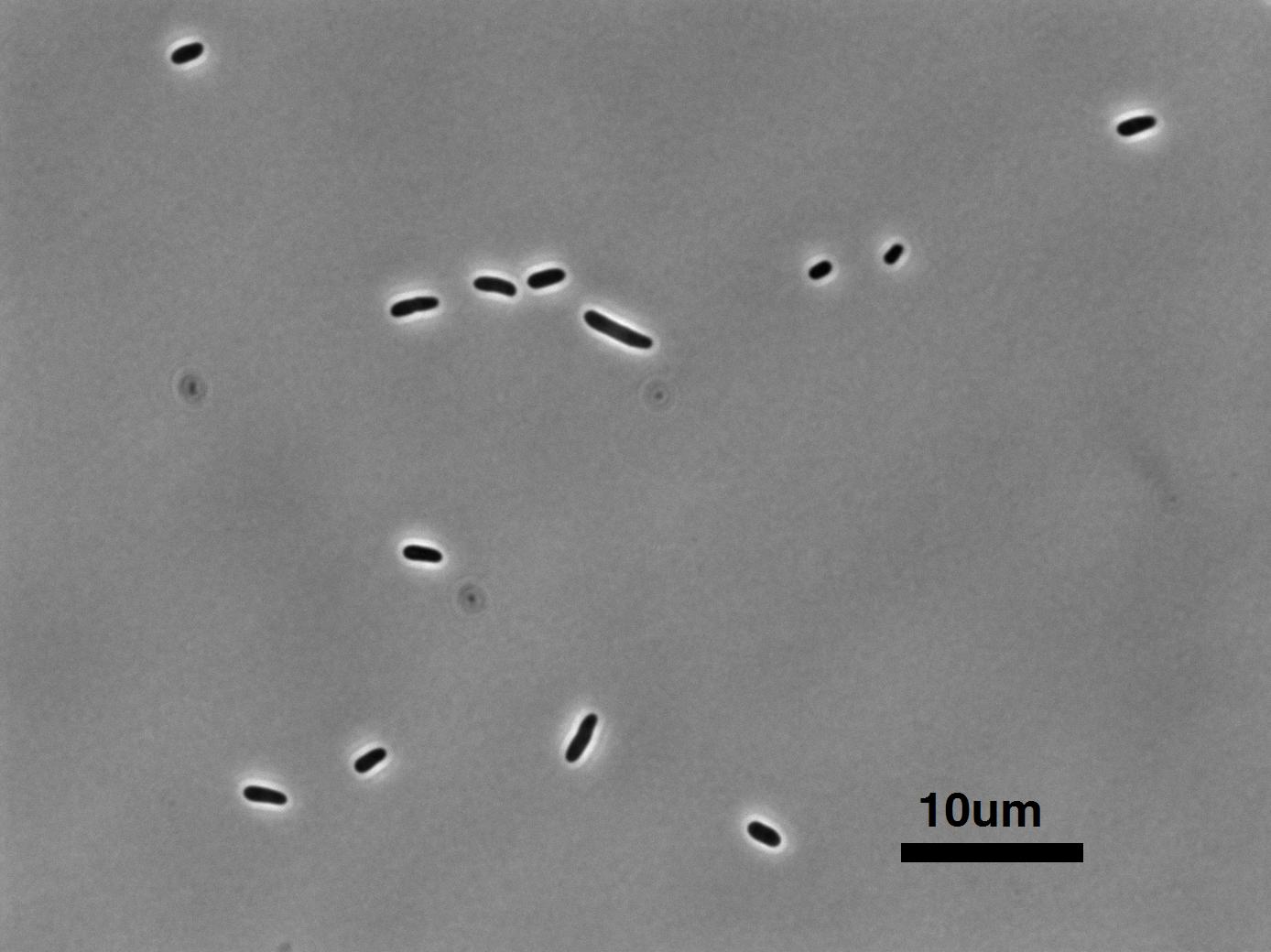
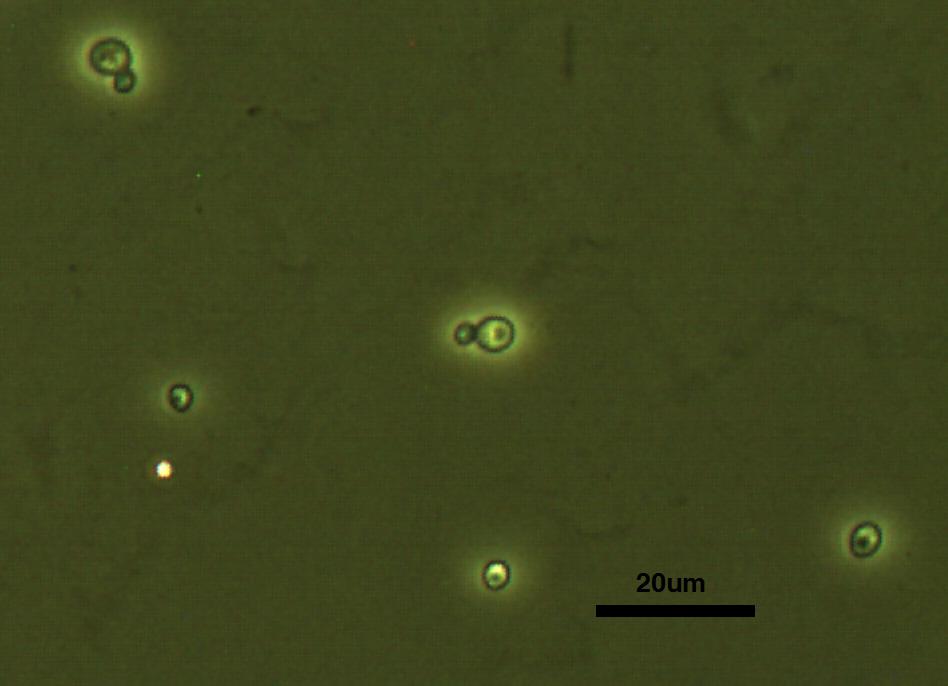
considering. In the prokaryotic setting the standard object is E.
coli to which all other prokaryotic cells can be compared;
likewise the yeast S. cerevisiae is the prototypical eukaryotic
cell. As such, after calibration the students always begin by
observing the size of these two organisms, with an emphasis on putting
scale bars on all of their pictures.
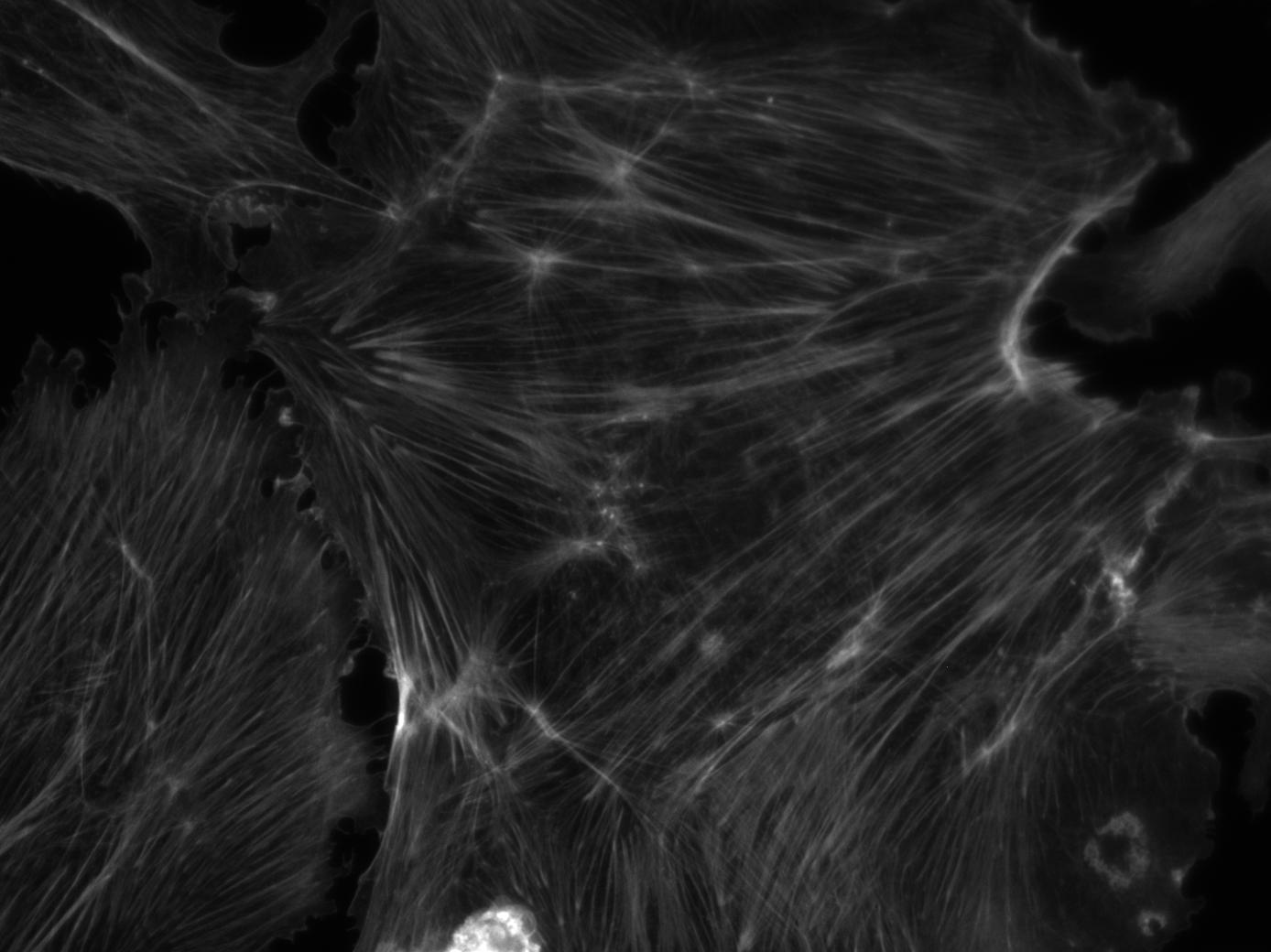
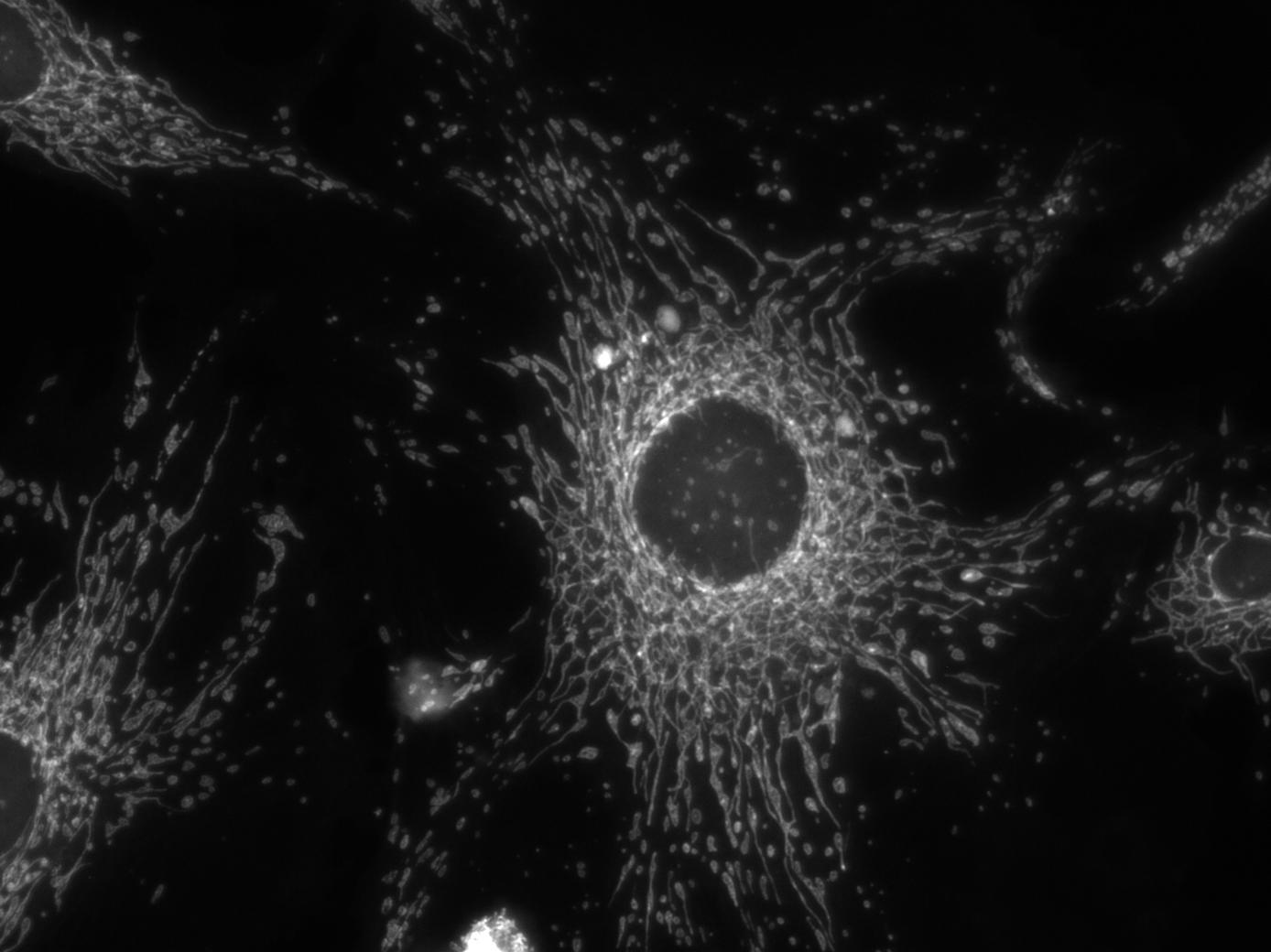
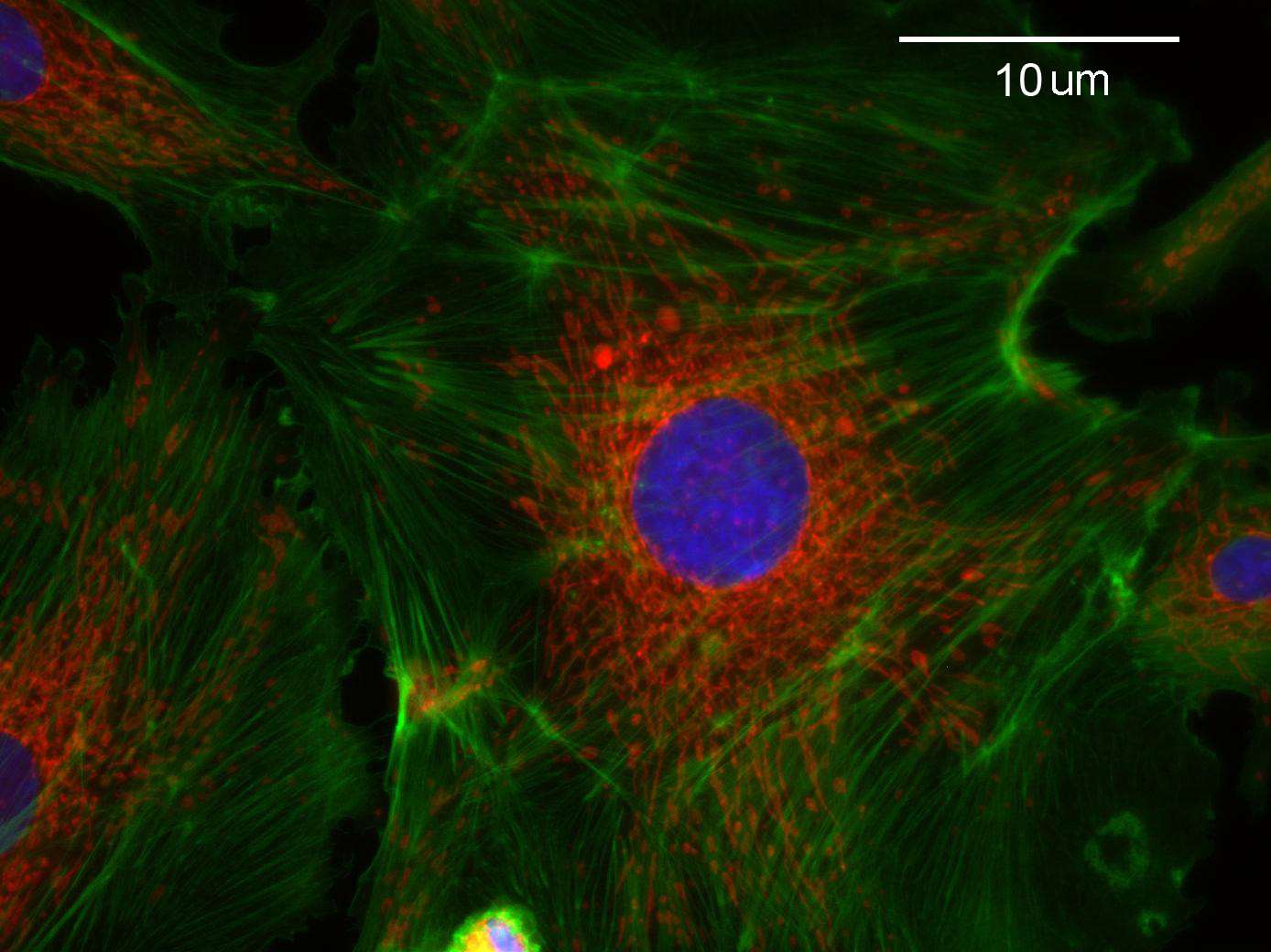
As
an exercise in fluorescence microscopy, the students take pictures
of bovine pulmonary cells in TRITC, FITC and DAPI fluorescence
modes to visualize the actin, mitochondria and nucleus respectively.
The students
then use Matlab(c) to combine the images into a fluoro-colored
image of the entire cell.
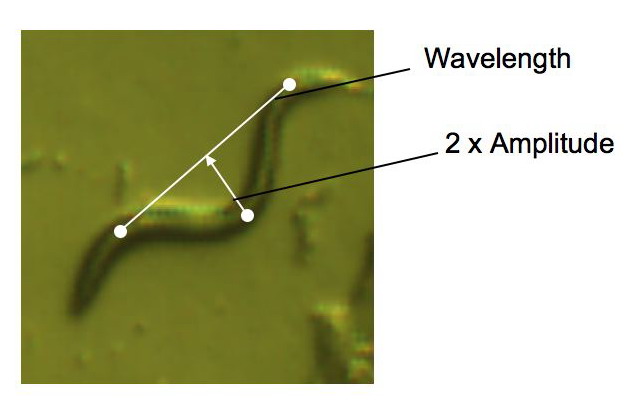
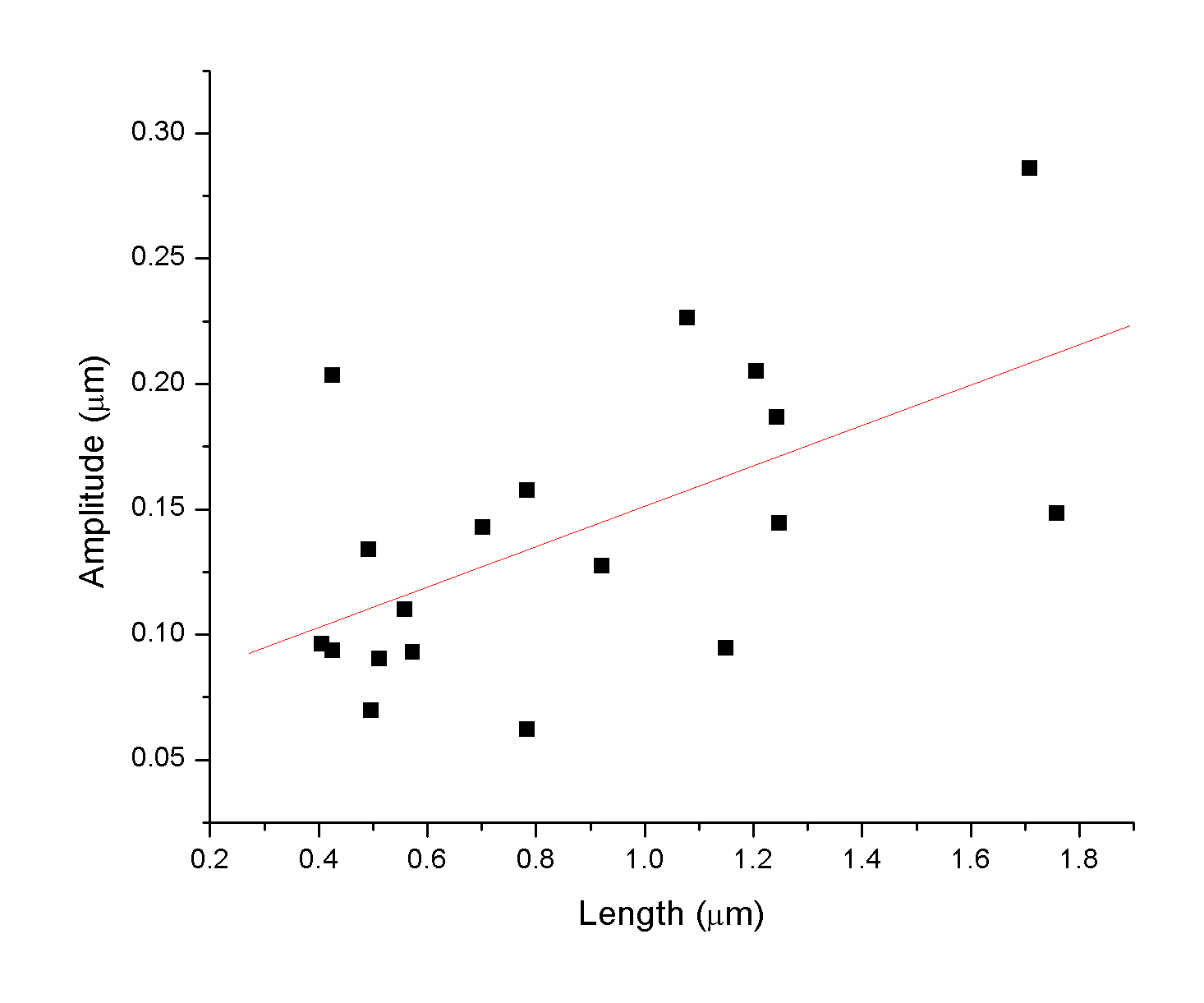
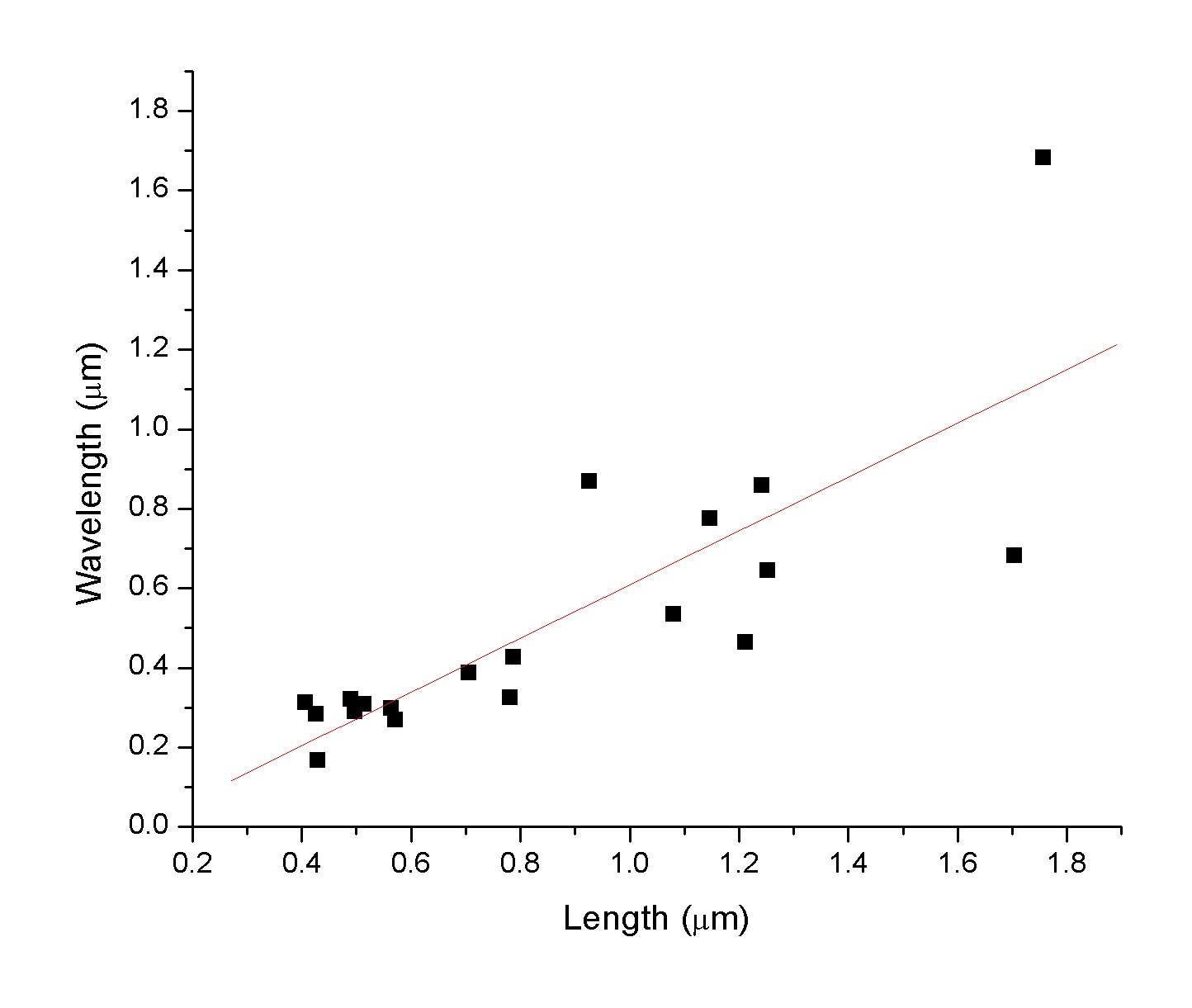
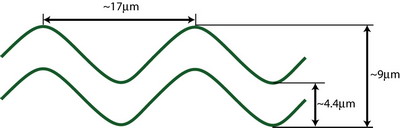
C. elegans is
a particulary important species in genetic studies. We simply like
to use them as dynamic organisms for understanding the size and
motility of multi-cellular organisms. Some students took the initiative
to measure the correlation between the geometric features of the
nematode (length and wavelength).
We became
interested in the algorithm employed by the nematode in its hunt
for food. Below is a real-time movie of a worm hunting around
the petri dish for food (E. coli).
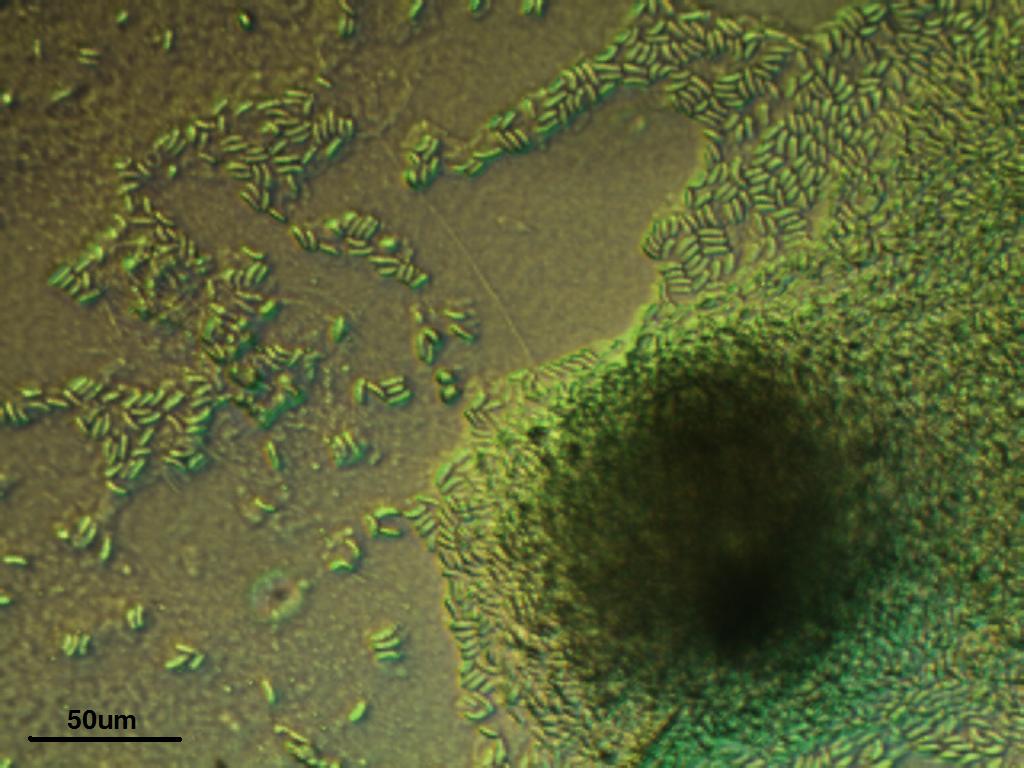
Nowadays
it's easy to order a wide array of organisms from a variety
of suppliers, but sometimes it's just fun to see what you have in
the
backyard. We took 1ml of mirky water from a pond on campus
and found a veritable zoo of interesting multi and single celled
organisms.
Here's a small sample:
Heliozoan -
notice about 1/3 of the way through the movie, some sort of cavity
closes on
the right-hand side. The dendritic structures are actually bundles
of actin filaments used for motility.
An unidentified
flagellate in the process of budding a new cell - flailing
in vain. We had considered putting this video to some
kind of dance beat.
This amazingly long
algae has a well defined cellular frequency. It was great to watch
it travese steadily across the field of view.
Stentor is
an incredibly large and dynamic single-celled organism, with
various organelles visible within the cell using simple brightfield
microscopy.
There are also fine, fast moving cilia around the "mouth" which
make amazing flow patterns (see The Rate of Things).
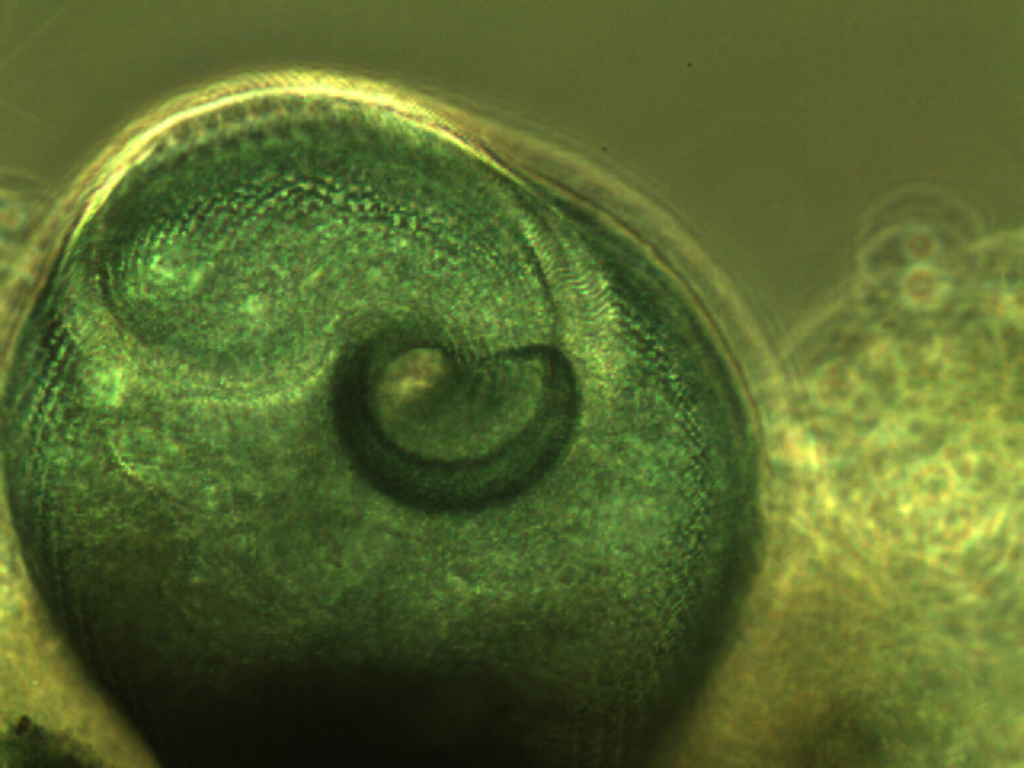
Dictyostelium
have an amazing life cycle, during which they sometimes
act as indepedent cellular organisms, but under the right conditions
will differentiate into fruiting bodies. Below are two great pictures
showing a fruiting body that has recently burst to release new, viable
cells. For 'slug' formation see The Rate of Things.
Spirulina is
another slow moving algae with an amazingly regular single-helix
structure. Using phase constrast microscopy, students characterized
this helix.